
Sigade aretus
Sigade aretuses on 20. sajandil hakatud järjest rohkem kasutama puhasaretuse asemel ristandaretust. Hübriidid on elujõulisemad. Sigade aretajad on vaatluse alla võtnud kõrge päritavusega tunnused nagu sigade kasvukiirus ja lihakeha tailiha sisaldus. Kasutades arvutitehnoloogiaid on kasutama hakatud ka madalama päritavusega tunnuste aretamist nagu pesakonna suurus ja põrsaste elujõulisus.
Sigade ristandaretuse programm Marmorliha on alguse saanud 1995 .aastal, millal imporditi Rootsist Eestisse hämpširi tõugu ja 1999.a. Austriast pieträäni tõugu emiseid ja kulte, et võimaldada kvaliteetse liha tootmist ristandaretuse baasil.
Eesti Tõusigade Aretusühistu (ETSAÜ) eesmärgiks on aretada ja levitada väärtuslikke tõusigu, et tagada sigade jõudlusvõime ja geneetilise väärtuse suurenemine ning seakasvatuse majanduslik tasuvus. Efektiivne ja jätkusuutlik seakasvatus nõuab nii kompetentsust kui ka erinevate standardite järgimist. ETSAÜ aretusprogramm taotleb lisaks aretuse juhtimisele suunata Eesti seakasvatust arvestama ka ühiskonna tervist, keskkonna ja loomade heaolu. ETSAÜ püüab kindlustada tasakaalustatud valikut ehk kompenseerida puudujääke ühes tunnuses mõne teise valikutunnuse kõrge tasemega. Selleks on kasutusele võetud aretusväärtused jõudlusele ja viljakusele.
ETSAÜ jätkusuutliku aretusprogrammi raames jälgitakse kuut valdkonda: loomade tervis ja heaolu, liha kvaliteet, efektiivne tootmine, keskkond, toiduohutus ja ühiskonna tervis ning geneetiline mitmekesisus.
Loe lähemalt Eesti Tõusigade Aretusühistu koduleheküljelt http://www.estpig.ee/
Seatõud
Seatõuge on maailmas palju. Ligikaudu sajal neist on praktiline tähtsus, kuid kohalikke, aborigeenseid ja hääbuvaid tõuge kokku arvates ulatub nende arv 400-ni.
Viljakus
Viljakusnäitajad – pesakonna suurus, pesakonnamass ja säilivus võõrutamisel – on suure majandusliku tähtsusega (Yen jt., 1987). M. Varley ja R. Stedman (1994) on määranud emise näitajate osatähtsuse pesakonna suurusele (joonis 1), kust nähtub emise isiklike omaduste oluline mõju viljakusele. Eksperiment eelduseks oli, et suure varieeruvusega isiklike tunnuste mõned algandmed võivad selgitada olulist osa sigade viljakuse üleüldisest variatsioonist.
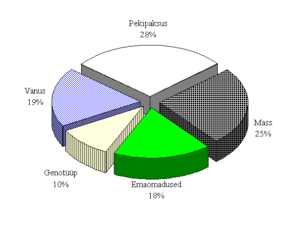
Joonis 1. Emise näitajate mõju pesakonna suurusele (Varley ja Stedman, 1994)
Üks tähtsamaid viljakust ja piimakust mõjutavaid tegureid on kindlasti põrsa sünnimass. L. Rydhmer (1992) leidis fenotüübilise ja geneetilise korrelatsiooni pesakonna suuruse ja põrsamassi vahel tiheda seose. Leiti, et pesakonna suurenemine on negatiivselt seotud põrsa-massiga. Suuremaid pesakond iseloomustab tendents, et sünnivad kergemad põrsad kui väiksemates pesakondades, niisiis võib aretus suurete pesakondade suunas viia suremuse suurenemiseni.
J.C. Kerr ja N.D. Cameron (1995) leidsid, et kui pesakonnas sünnib üks põrsas rohkem väheneb põrsaste võõrutuseelne suremus proportsionaalselt 0,12 põrsa võrra kuni põrsaste arv pesakonnas võrdub viiega, seejärel aga pesakonna suurenedes ühe põrsa võrra tõuseb ka põrsaste suremus proportsionaalselt 0,01 põrsa võrra (joonis 2).
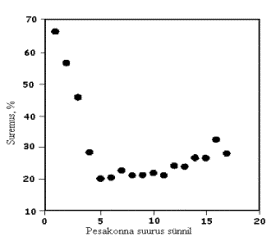
Joonis 2. Põrsaste võõrutuseelse suremuse sõltuvus pesakonna suurusest (Kerr ja Cameron, 1995)
Väikestes pesakondades sündinud põrsad on suuremad, tugevamad ja elujõulisemad (Rydhmer, 1992; 1993). Nad haigestuvad vähem ja kasvavad kiiremini, mistõttu eriti suurte (nõrkade põrsastega) pesakondade tootmine ei ole majanduslikult õigustatud. Tootmisfarmides võiks eesmärgiks seada 10…11 põrsast pesakonnas, aasta keskmisena peaks emiselt saama vähemalt 20 põrsast (Meisner, 1990; Kersten, 1995).
J.C. Kerr ja N.D. Cameron (1995) on oma katsete põhjal koostanud graafiku põrsaste surevuse sõltuvuse kohta sünnimassist (joonis 3). Põrsaste suremus väheneb kiiresti põrsa sünnimassi suurenedes. Põrsamassi korral üle 1,5 kg on suremus on alla 10%.
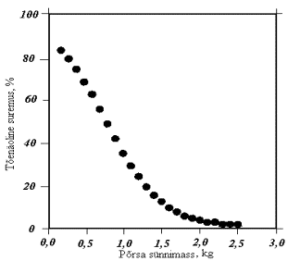
Joonis 3. Põrsaste võõrutuseelse suremuse sõltuvus sünnimassist (Kerr ja Cameron, 1995)
Oma töös on M.H. Fahamy ja C. Bernard (1971) põhjalikult käsitlenud pesakonna suuruse ja põrsaste elujõulisuse seost. Elusaltsündinud põrsaste suur arv ei taga veel, et suur arv põrsaid ka võõrutatakse. Põrsa sünnimass on aga elujõulisusega positiivselt seotud just esimeste nädalate kestel. Seega tuleb põrsaste arvu kõrval arvestada ka põrsaste sünnimassiga.
Ka H.E. Nielsen (1975) väidab, et suuremas pesakonnas väheneb tugevasti põrsaste elujõulisus põrsaste väiksema massi tõttu. Põrsaste suremus on kõige suurem siis, kui põrsa sünnimass on alla 1,20 kg.
Vaatamata pesakonna suuruse negatiivsele seosele teiste viljakusnäitajatega on üheks aretuseesmärgiks suurte pesakondade saamine võõrutamisel. Elusaltsündinud põrsaste arv on üheks selektsiooni programmi valikutunnuseks Rootsis ja paljudes teistes maades (Brandt, 1990; Webb, 1991; Estany jt., 1993; Rydhmer, 1993; Sehested ja Ianssin, 1993). Sündinud põrsaste arvu asemel võõrutatud põrsaste arvu valiku aluseks võtmast takistab arvatavasti suhtumine, et sündinud põrsaste arv sõltub emisest, hiljem aga mõjutab ka farmer põrsaste säilivust. Et põrsad on esimeste elunädalate jooksul väga tundlikud keskkonnatingimuste muutuste suhtes, siis peab farmer tagama soodsad tingimused põrsaste säilimiseks ja arenguks. Põrsaste arvu kasvuga pesakonnas suureneb ka suremus, seega on geneetiline edu vähe väärt, kui see ei kajastu võõrutatud põrsaste arvu suurenemises. Seepärast on pesakonna suurus võõrutamisel parem valikutunnus kui pesakonna suurus sünnil, sest parem põrsaste säilivus näitab nende suuremat elujõulisust antud tingimustes. Siiski, kui kasutatakse ristandeid ja võõrutusvanus varieerub, on raske kehtestada standardit pesakonna suurusele võõrutamisel (Rydhmer, 1993).
Ristamise mõju hindamisel sigade viljakusele tuleb arvestada võrreldavate rühmade suurust. Katsetes on sigade arv sageli väike, mis ei võimalda teha objektiivseid järeldusi uurimistulemustest. Pig Improvement Company (1983) poolt on esitatud pesakondade miinimumarvud ja keskmiste 95% tõenäosusega erinevused (tabel).
Nendest nähtub, et objektiivsete järelduste tegemiseks peab pesakondade arv võrreldavates rühmades olema küllaltki suur. Siit võib ka selgust tuua küsimusele, miks mõned katsetulemused ei ühti teiste katsetulemustega.
Tabel. Pesakondade miinimumarvud ja keskmiste erinevus (Pig Improvenent Company, 1983)
| Pesakondade arv kummaski rühmas | Rühmade keskmiste erinevus, korda |
| 25 | 1,58 |
| 50 | 1,02 |
| 100 | 0,79 |
| 150 | 0,64 |
| 250 | 0,50 |
| 500 | 0,35 |
Väikeste rühmadega saab kergemalt hinnata siis, kui need on läbi viidud mitmes korduses ja tulemused on ühesuunalised. Edaspidi tuleks sigade viljakust käsitlevates uurimustes suuremat tähelepanu pöörata võrreldavate rühmade suurusele.
Pärilikud tegurid
Seakasvatuse efektiivsust saab parandada kahel viisil:
- parandades genotüüpi;
- parandades keskkonda ehk mittegeneetilisi tegureid.
Genotüüp seab lae, mil määral fenotüüpi saab parandada keskkonnategurite, pidamise ja söötmise parandamisega, aga alati on tähtis taotleda sigade geneetilise võime parandamist (English jt., 1988).
Paljud uurijad on püüdnud kindlaks teha geneetilise edu puudumise põhjusi pesakonna suuruses nii aretuskarjades kui ka valikueksperimentides (Skjervold, 1979; Johansson, 1981; Southwood ja Kennedy, 1991; Bolet jt., 1989). Mitmed kaasaegsed uurimistulemused, mis baseerusid viljakate emiste pesakonna andmetele, näitasid, et valik suuremate pesakondade järgi võib olla edukas (Legault, 1985; Bichard ja David, 1985). Tänapäeva teadlased on optimistlikud pesakonna suuruse geneetilise potentsiaali suhtes (Avalos ja Smith, 1987; Haley jt., 1988). A.J. Webb (1991) konstateerib, et on praktiline tegelikkus geneetiliselt suurendada pesakonda aastas 0,2…0,3 põrsa võrra.
Väike geneetiline edu viljakuses, eriti pesakonna suuruses, on saavutatud kaasajal (Kaplon jt., 1991; Southwood ja Kennedy, 1991). Võrreldes metssigadega on toimunud oluline muutus (Aumaitre jt., 1982). Väikese valiku edu põhjuseks pesakonna suuruses pole variatsiooni madal tase, sest tõugudevaheline geneetiline variatsioon on suur. Hiinast pärit meishani tõu kõrge viljakus on potentsiaali sobivaks näiteks, kuid pesakonna suuruse päritavus on sellest hoolimata madal (Bidanel, 1988). 1970. aastatel tõstatasid G.S. Strang ja J.W.B. King (1970) küsimuse pesakonna suuruse aretusväärtusest. Sellel ajal põhines peaaegu kogu valik toodangunäitajatel, peamiselt massi-iibel ja tailiha osatähtsusel. Nende tunnuste osas on saavutatud suurt edu (Lundeheim ja Eriksson, 1984) ja paljud uurijad eeldavad et tailiha optimum saavutatakse peatselt (Ollivier jt., 1990). Pealegi on selektsioon sega-mudelitega suurendanud lootust, et geneetilist edu võib saavutada tunnuste osas, millel on madal päritavus ja mida on mõjutavad rohkem keskkonna tingimused (Long jt., 1991). Pesakonna suurust on käsitletud näitajana, millel on suur majanduslik väärtus (Legault, 1978; Smith jt., 1983). See on samuti ainus viljakusnäitaja, mis on kantud paljudesse aretusprogrammidesse (Rydhmer, 1993). Tänapäeval on paljud aretusfirmad ja tõuraamatud seadnud prioriteediks suurendada viljakusnäitajate osa valikul.
Ebasoodne geneetiline korrelatsioon viljakuse ja teiste valikus kasutatavate näitajate vahel, keskkonna mõju ja nõrk geneetiline korrelatsioon esimese pesakonna viljakuse ning eluaja viljakuse vahel võivad olla viljakuse tähtsust vähendavad faktorid. Enamik teadlasi on seisukohal, et otstarbekam on parandada viljakusnäitajaid ristamiskombinatsioonidega, saavutades edu soovitud tunnustel ja täiendavalt manipuleerida keskkonnategurite ning majandamisega. Selline valiku ja pidamise koosmõju arvestamine kehtib ka lihakehanäitajate parandamisel (Johansson, 1981). Näiteks leidsid G. Bolet jt. (1984) negatiivse geneetilise seose lihasuse ja pesakonna suuruse vahel, mis on üks viljakust vähendavaid tegureid. L. Rydhmer (1993) seevastu leidis soodsa geneetilise korrelatsiooni lihasuse ja pesakonna suuruse vahel. Ka O. Vangen (1980) leidis, et valik suure toodangu suunas võib olla seotud pesakonna suuruse väikese kasvuga sünnil. Kontrastiks sellele leidsid K. Johansson ja B.W. Kennedy (1983) ebasoodsa geneetilise korrelatsiooni pekipaksuse ja pesakonna suuruse vahel. Vastupidiselt neile leidis L. Rydhmer (1993) ebasoodsa geneetilise korrelatsiooni kasvukiiruse ja pesakonna suuruse vahel. D.J.A. Cole (1971) andmetel pole aga ilmnenud tõendeid negatiivsete geneetiliste seoste kohta viljakuse ja teiste majanduslikult kasulike tunnuste vahel. P.W. Ferguson jt. (1985) leidsid positiivse geneetilise korrelatsiooni emise kehamassi ja pesakonna suuruse vahel. Soodsa korrelatsiooni massi-iibe ja 3-nädalase pesakonna suuruse vahel leidsid K. Johansson ja B.W. Kennedy (1983), ehkki see oli nõrgem võrreldes pesakonna suurusega sünnil. Põrsamass pole positiivselt seotud mitte ainult elujõulisusega, vaid kogu eluaja jõudlusega (Rydhmer, 1993). Geneetiline varieeruvus põrsa sünnimassis on suur. Järelikult näib kasulik olevat seada ka see aretuseesmärgiks.
L.D. Young jt. (1976) leidsid uurimuses soodsa geneetilise korrelatsiooni toodangunäitajate ja põrsamassi vahel. Emiste valikul kõrge toodanguindeksi järgi suureneb põrsa sünnimass (Vangen, 1980) ning saadakse suuremad sead (Jacques ja Vangen, 1987). R.M. McKay (1990) võttis valiku aluseks suure massi-iibe ja lihasuse seitsmes põlvkonnas, aga ei leidnud olulist seost emise kehamassi ja pesakonna suuruse vahel. On tõsiasi, et ühest viljakama tõu – meishani emised on üsna väikesed, mis näitab, et suur kasv ei ole suurte pesakondade saamiseks vajalik (Bidanel jt., 1990). Seoseid massi-iibe, keha mõõtmete ja pesakonna suuruse vahel on üldistavalt käsitlenud F.D. Brien (1986), märkides, et tulemused sigadel olid vastuolulised. K. Johansson (1981) ja L. Rydhmer (1993) on seisukohal, et vastuolusid erinevates uurimustes viljakuse ja toodangunäitajate vahel peab käsitlema ettevaatlikult, sest tunnuste definitsioonid ja keskkonnatingimused erinevad uurimustes suuresti.
Seakasvatusettevõtetes kasutatakse sageli suurema kasumi saamiseks heteroosi, sest ristandloomade jõudlus ületab nende puhtatõuliste kaaslaste keskmisi näitajaid. Tõugude ristamisel tekib heteroos, kui ristandite jõudlus on suurem puhtatõuliste vanemate keskmisest (joonis 1). Ristandite jõudlus võib olla suurem või väiksem kui parima puhtatõulise vanema oma, sõltudes ristatavatest tõugudest (Clutter jt., 1990).
Kunstlik seemendus
Kui on kindlaks tehtud, et emisel on kuldi suhtes paigalseisurefleks ja kui seda saab esile kutsuda ka inimese poolt, tuleb hakata tegema ettevalmistusi emiste kunstlikuks seemenduseks:
- Spermatuub võetakse säilituspakendist välja, raputatakse ettevaatlikult ja soojendatakse kuni 38 °C.
- Seemendamisel tuleb kaasa võtta seemenduskateeter, käärid, parafiin ja puhas paber. Kateetreid tuleb säilitada nii, et nad oleksid kasutamisel puhtad ja tolmuvabad.
- Emise paigalseisurefleksi esilekutsumine.
Kõrvaldada tuleb segavad faktorid: emise tähelepanu ei tohi kõrvale pöörata (sööt), teda ei tohi rahutuks ajada (lärm, võõrad isikud) või hirmutada (toores kohtlemine); seemendust peab läbi viima inimene, kellega loomad on harjunud. Seemendatav emis tuleks enne kunstlikku seemendust puhastada.
Emise stimuleerimine kuldi poolt: seemendatav emis eraldatakse teistest emistest ja viiakse kuldi lähedusse. Sel teel on võimalik pilk-, kuulmis- ja haistmiskontakt kuldiga läbi suluvõrestiku. Samuti võib lasta kuldil läbi vahekäigu emiste eest mööda joosta.
Emise stimuleerimine inimese poolt: pärast esimest ettevaatlikku kontakti stimuleeritakse emise paigalseisurefleksi külje- ja landehaarde abil ning ratsatestiga. Seda võib teha seemendaja ise või keegi abiline.
Paigalseisurefleksi esilekutsumiseks võib mängida emistele helilinti (kuldi paaritusaegsed ruiged) või pihustada kuldi lõhna.
Emakakaelakanali vahelduva rütmilise kokkutõmbumise ja lõtvumise tõttu transporditakse sperma aegamööda emakasse. See protsess võib kesta 10-15 minutit. Tugevasti väljenduva inna ja seemendamisel esilekutsutud paigalseisurefleksi puhul toimub kontraktsioon sageli kiiremini ja sperma “imetakse” tihti juba umbes 5 minuti jooksul emaka poolt sisse.
Seemenduse ajal peab kateeter olema õige asendi puhul emakakaelakanalis sedavõrd tihedalt, et sperma voolaks emaka suunas ja mitte tagasi tuppe. Pipetid on seemendusseadmed, mis on ette nähtud kas korduvaks (kummipipetid) või ühekordseks kasutuseks (plastmassist pipetid). Hügieenilistel ja majanduslikel põhjustel kasutatakse tänapäeval enamasti plastmassist pipette, mis pärast kasutamist hävitatakse. Neid seemenduskateetreid saab osta seemendusjaamadest. Kasutatavad pipetid on 40-50 cm pikkused. Nad erinevad üksteisest oluliselt oma otsiku kuju poolest. Viimased võivad olla erineva suurusega, ovaalse kuni peaaegu täisnurkse paksendiga või varustatud korgitsalaadse vasakule pööratud vindiga.
Viimasena nimetatud kateetrimudel sarnaneb kuldi peenisega ja see keeratakse seemendusel emakakaela. Mugava käsitsemise tõttu kasutatakse taolist kateetrit sageli.
Kunstliku seemenduse läbiviimine:
- Häbememokad tuleb vajadusel puhastada kuiva puhta paberiga, et vältida mustuse sattumist tuppe.
- Pipetiotsik tehakse mõne parafiinitilga või eelnevalt lahtilõigatud tuubist võetud spermaga libedaks.
- Häbememokad aetakse laiali pöidla ja nimetissõrmega, nii et kateeter oleks kontaktis ainult tupeesiku limaskestaga.
- Seemenduskateeter lükatakse nüüd altpoolt, umbes 450 nurga all tupe suunas tupeesikusse. Kui ligikaudu 10 cm pipetist on sisse viidud, tõstetakse tema otsa veidi üles ja lükatakse pipetti edasi horisontaalses (rõhtsas) asendis mööda tupevõlvi. Sel viisil ei satuta kusejuha avausse, mis asub tupepõhjal.
- Ligikaudu 20-25 cm järel on tunda vastupanu, mis tuleneb emakakaela kurdudest.
- Nüüd püütakse tundliku vajutusega kateetrit nii kaugele kui võimalik emakakaela kurdudesse sisse keerata.
- Kui edasi ei ole enam võimalik keerata, on pipetiotsik piisavalt kindlalt paigal.
Seemendamine
- Seemenduspipeti otsik painutatakse ülespoole ja soojendatud spermatuub paigutatakse horisontaalselt ning hoitakse nii kogu seemenduse vältel. Kateetri õige asendi korral hakkab sperma emaka rütmiliste imemisliigutuste tõttu aeglaselt spermatuubist välja voolama.
- Kerge surve seemendustuubile toetab seda protsessi.
- Sigade edukaks viljastamiseks on spermide arvu ja kvaliteedi kõrval väga tähtsal kohal ka kogu spermaportsjoni maht. Seetõttu tuleb jälgida, et alati kasutataks ära terve tuubi sisu. Seemendusprotsessi võib pidada lõppenuks alles siis, kui ka kateetri sisu on jõudnud emakasse. Seejärel eemaldatakse pipett sisseviimisele vastupidises suunas ettevaatlikult keerates emakakaelast ja tõmmatakse välja.
Raskused sperma sisseviimisel võivad olla põhjustatud kateetri valest asendist või spermatuubi liiga tugevast pigistamisest. Kui sperma ei voola üldse välja, voolab tagasi tuubi või tupest välja, tuleb korrigeerida pipeti asendit. Kui seemendusel kasutatakse liiga tugevat survet, voolab sperma emaka asemel tuppe. Kui emis muutub seemenduse ajal rahutuks või veritseb, võib see olla põhjustatud tupepiirkonna või kusejuha vigastustest.
A. Lember, V. Luts, Ü. Roosmaa, A. Oja. Eesti Maaülikool, 2014
Seemenduse nõuded
| Vara indlev emis | pikk innaperiood |
| Inna algus: | 3/4. päev pärast võõrutamist |
| Inna kestus: | 3 päeva (või kauem) |
| Seemendamise aeg: | 1.kord=24-36 tundi pärast inna algust* 2.kord=12-16 tundi pärast 1.seemendust 3.kord=12-16 tundi pärast 2.seemendust *ainult väga pika innaperioodi puhul |
| Normaalselt indlev emis | |
| Inna algus: | 5.päev pärast võõrutamist |
| Inna kestus: | 2,5 päeva |
| Seemendusaeg: | 1.kord=12-24 tundi pärast inna algust* 2.kord=12 tundi pärast 1.seemendust |
| Hilisev indlev emis | lühike innaperiood |
| Inna algus: | 6/7 päev pärast võõrutamist |
| Inna kestus: | 2 päeva (või vähem) |
| Seemendusaeg | 1.kord=12 tundi pärast inna algust* 2.kord=12 tundi pärast 1.seemendust, kui emisel on veel paigalseisurefleksi *Soovitatud paaritus- ja semendusajad kehtivad ainult siis, kui innakontrolli viiakse läbi kaks korda päevas |
Teoreetiliselt võib ka ühekordse paarituse või seemendusega saavutada häid viljastamistulemusi. Eelduseks on õige paaritus- või seemendusaeg. Isegi kaks korda päevas läbiviidava innakontrolli puhul, kui tehakse kindlaks täpne innaperioodi algus, on keeruline ovulatsiooni aega täpselt prognoosida. Praktikas on seetõttu soovitav kasutada topeltseemendust või -paaritust.
Inna stimuleerimine
Inna stimuleerimisel on väga tähtis kontakt kuldiga haistmis-, kuulmis-, nägemis- ja kompimisorganite kaudu. Tänapäeva kogemusi arvestades ei tohiks emisepidaja loobuda paarituslaudast. Ümberpaigutamine paarituslauta ei tähenda võõrutatud emiste ja nooremiste jaoks mitte ainult kontakti kuldiga, vaid ka uusi keskkonnamõjusid. See stimuleerib ja aitab emistel kergemini inda esile kutsuda. Ka enamasti kunstlikku seemendust kasutavad ettevõtted ei tohiks katsukuldi kasutamisest loobuda.
Kui emistel on indlemisega probleeme, võib sellel olla mitmeid põhjusi. Emisepidaja peaks siis kontrollima järgmist.
- Kas paarituslaudas on piisavalt valgust? Kui valgust on liiga vähe, võib olla abi kunstliku valguse (12 tundi ööpäevas) kasutamisest. Loomalaudas peaks olema sama valge kui büroos.
- Kas kult stimuleerib emiseid optimaalselt? Noored kuldid stimuleerivad hästi alles 10 kuu vanuselt. Seetõttu ei ole kasulik vana kulti enne välja vahetada, kui noor kult piisavalt kogenud on.
- Kas paarituslaut on hästi isoleeritud ja piisavalt õhutatud? Talvine niiskus ja külm ning suvine kuumus võivad olla inna puudumise põhjuseks. Lisasoojusallikas paarituslaudas aitab vältida suuri päevaseid ja öiseid temperatuurikõikumisi. Suvine suur kuumus tekitab loidust. Selle tagajärjeks on ebapiisav indlemine, ümberindluse suur osakaal ja väikesed pesakonnad.
- Kas emisel on piisavalt liikumisvabadust ja sotsiaalseid kontakte? Nooremiseid tuleks hoida gruppides. Liiga varane lõastamine või puuri panemine mõjuvad ebasoodsalt. Ka võõrutatud emistele mõjub liikumine inda stimuleerivalt.
- Kas emist söödetakse õigesti? Liiga suur massikadu imetamisperioodil, eriti pärast esimest pesakonda, või nooremiste ülesöötmine avaldavad viljastumisele ebasoodsat mõju. Mitte mingil juhul ei tohi anda emistele riknenud sööta.
- Kas nooremised on piisavalt vanad ja täielikult karja integreerunud? Nooremiseid ostetakse karja enamasti 180 päeva vanuselt. Nn. transpordi-inna ajal ei tohiks nooremist veel seemendada. Selle kohta tuleks teha hoopis märkmeid ja oodata seejärel järgmist või ülejärgmist innaperioodi. Nooremis peaks olema esmasel paaritamisel vähemalt 210 päeva vanune ja kaaluma 110 kg.
- Kas emised on terved? Põletikud, sõravigastused või muud haigused on sageli inna puudumise põhjused. Õigeaegse ravi ja järjekindla vaktsineerimise abil saab hoida karja hea tervise juures. Siinkohal tuleks regulaarselt konsulteerida loomaarstiga. Kui haigussümptomid korduvad, tuleks otsida põhimõttelisi vigu loomade karja toomise, karja majandamise ja hügieeni tasandil.
Üksikjuhtudel on mitteindlemise puhul mõttekas kasutada hormoonravi. Kui nn. mitteindlejal käib siiski juba korrapärane tsükkel ja inda ei avastatud õigeaegselt, on oluline, et hormoonravi toimuks tsükli jooksul õigel ajal.
Probleememiste hormoonravi võib vajadusel läbi viia ainult loomaarsti järelevalve all. Hormoonravi ei ole mitte ainult väga kallis, vaid enamasti pole vastava sisustuse ja töökorras paarituslaudaga farmis selle järele vajadust.
Näpunäiteid innakontrolliks:
- Kes ei soovi innakontrolli ja seemendusi läbi viia nädalavahetusel, peaks võõrutama neljapäeval või reedel.
- 3. päeval pärast võõrutamist peaks esimest korda kontrollima, kas loom ei indle, et teha kindlaks enneaegsed indlejad. Mõnes farmis võetakse raskemad põrsad mõni päev enne õiget võõrutuspäeva emise juurest ära. Sellistel emistel algab innaaeg sageli varem.
- Indlemist tuleks põhimõtteliselt kontrollida 2 korda päevas. Üks kord päevas kontrollides ei ole võimalik täpselt kindlaks teha paigalseisurefleksi esinemist.
- Innakontrolli on kõige parem läbi viia siis, kui ajada emised kuldi juurde või vastupidi. Kartlikud ja väga uudishimulikud loomad peaksid jääma harjumuspärasesse keskkonda ja paarituspartner tuleks nende juurde tuua. Kui kulti pole, on abi kuldilõhna kasutamisest.
- Inna kontrollimine peaks toimuma laudas “vaiksel tunnil”. Mingil juhul ei tohiks kontrollida emiseid söötmisaegadel. Loomadega tegelemisel peaks tingimata vältima närvilisust, kuna muidu võib juhtuda, et paigalseisurefleks tuleb esile liiga hilja ja indlevat looma ei tunta õigel ajal ära. Inna algus ja pikkus on ühe looma puhul tihti korduvad. Seetõttu on selliste andmete jäädvustamisest hilisemate seemenduste puhul palju kasu.
Näpunäited kunstlikuks seemenduseks ja sperma säilitamiseks:
- Paljudes ettevõtetes viiakse emis kuldi sulu ette ja paigalseisurefleksi tekkides seemendatakse teda kunstlikult. On väga tähtis, et seemendus viidaks läbi sobival ajal ning seda tehtaks rahulikult.
- Kasutada ainult puhtaid, tolmuvabu, ühekordseks või mitmekordseks kasutamiseks ettenähtud pipette.
- Häbet puhastada kuivalt, kui vaja, siis tselluloospaberiga.
- Enne seemendamist kutsuda esile paigalseisurefleks landehaarde, ratsatesti või paaritamiskoti abil. Pipett teha libedaks ainult parafiinõli või sperma abil, kuna muud ained võivad spermat kahjustada.
- Pipett tuleb viia sisse põiki altpoolt ülespoole kuni emakakaelakanali keskosani. Emis peab pipetti “kinni hoidma”, s.t. pipeti tagasitõmbamisel peab olema tunda kerget vastupanu.
- Tuubi otsa eemaldamisel ja tuubi pipetile asetades peab sperma voolama ilma surveta või ainult kerge survega emise suguorganitesse.
- Seemendus peaks kestma kauem kui 3 minutit ja emist tuleks enne ja pärast seemendust ning seemenduse ajal stimuleerida.
- Kui spermatuub on tühi, ei tohiks pipetti kohe välja tõmmata. See väldib sperma tagasivoolu.
- Sperma säilitamine. Kunstliku seemenduse õige läbiviimise kõrval on oluline sperma nõuetekohane säilitamine ettevõttes.
- Sperma säilitamistemperatuur on 16 kuni 18o C. Eriti kahjulikult mõjuvad spermale madalamad temperatuurid (alla 12o C). Kõrgete temperatuuride puhul (üle 20o C) säilivusaeg väheneb. Vältida tuleb tugevaid temperatuurikõikumisi.
- Spermat tuleb kaitsta päikesekiirte (UV-kiirte) eest.
- Spermadoose tuleks kaks korda päevas loksutada, et spermid oleksid toitelahuses hästi segunenud. Optimaalsetes tingimustes säilitamisel püsib sperma viljastamisvõime ühtlaselt kolme päeva jooksul.
A. Lember, V. Luts, Ü. Roosmaa, A. Oja. Eesti Maaülikool, 2014
Süstimistehnika
Sigade süstimiseks kasutatakse mitmesuguse suurusega süstlaid ja nende juurde kuuluvaid süstlanõelu e. kanüüle. Nõela ühendamiseks süstlaga võib vahetükina kasutada peenikest kummivoolikut, mis väldib nõela murdumist. Süstlad ja nõelad steriliseeritakse enne süstimist keetmisega. Kui süstal ja nõelad ei ole mäda ning mustusega saastunud, võib neid desinfitseerida ka 70° alkoholiga.
Naha alla süstitakse neisse piirkondadesse, kus alusnahk on hästi arenenud ning nahk seetõttu hästi liikuv ja kergesti volti tõstetav. Sigadele tehakse nahaalust süstimist tavaliselt kõrva taha või kubemevolti. Enne protseduuri tuleb süstimiskoht desinfitseerida (70° alkohol, joodi/piirituslahus). Süstimisel tõstetakse vasaku käe pöidla ja nimetissõrme abil nahavolt üles ja viiakse parema käega ainult nõel või süstla otsa asetatud nõel voldi jalamilt naha alla umbes pooles pikkuses. Kui nõel on naha all, siis ta ots liigub vabalt. Süstimine toimub kergesti ja süstekohale tekib kubel.
Lihasesse süstitud ravimilahused imenduvad aeglasemalt kui nahaalusest sidekoest. Seetõttu on lihasesse süstitud ravimite toimeaeg pikem. Lihasesse süstimine on vähem valus kui süstimine naha alla. Süste tehakse tavaliselt paksu lihaste kihiga piirkonda, nagu abapiirkonda ja kaela. Vanematele sigadele lihastesse harilikult ei süstita, sest paksu nahaaluse rasvakihi tõttu võib süstitud lahus kergesti sattuda pekisse, kust ta aga ei imendu ega avalda toimet. Süstimisel viiakse nõel pärast torkekoha ettevalmistamist järsu tõukega risti läbi naha lihasesse, millele järgneb ravimilahuse sissesurumine.
Süstimine veeni toimub sigadel tavaliselt kõrvalesta välisküljel paiknevatesse veenidesse. Süstimisel sea kõrvaveeni tõmmatakse kõrvajuure ümber nöör või kummivoolik, mille tagajärjel veenid kõrvalestas täituvad. Algul süstekoht desinfitseeritakse, nõel viiakse läbi naha mõnesse suuremasse veeni ja hoitakse kinni koos kõrvalestaga. Kaltsiumkloriidi, kloraathüdraadi jt. ärritavate ravimite manustamisel peab hoolikalt jälgima, et süstitavat lahust ei satuks veeni ümbruse kudedesse (tekivad kudede kärbus ja veenipõletik). Selleks tulebki nõel süstimisel pikalt veeni viia, et ta looma rabelemisel kergesti välja ei tuleks. Pärast süstimist veen komprimeeritakse (surutakse kokku), lastakse verd läbi nõela välja voolata. Seejärel lõpetatakse komprimeerimine, haaratakse nahk nõela ümber pöidla ja nimetissõrme vahele, surutakse vastu kõrva ja tõmmatakse nõel välja. Pärast nõela väljatõmbamist torkekoht desinfitseeritakse.
Süstimine kõhuõõnde on levinuim põrsastele ja kesikutele ravimite manustamise viis. Kõhuõõnde süstimisel hoitakse põrsad ja kesikud tagajäsemest rippu, pea allapoole. Pärast süstekoha desinfitseerimist viiakse nõel lühikese torkega häbemeluu ja naba vahe keskkohalt (tagapool naba) 1-2 cm kaugusel keskjoonest läbi kõhuseina ning tehakse süste.
A. Lember, V. Luts, Ü. Roosmaa, A. Oja. 2014